

The Lymphatic Endothelial Cell Secretome Inhibits Osteoblast Differentiation and Bone Formation
 , ,
, ,
Abstract
:1. Introduction
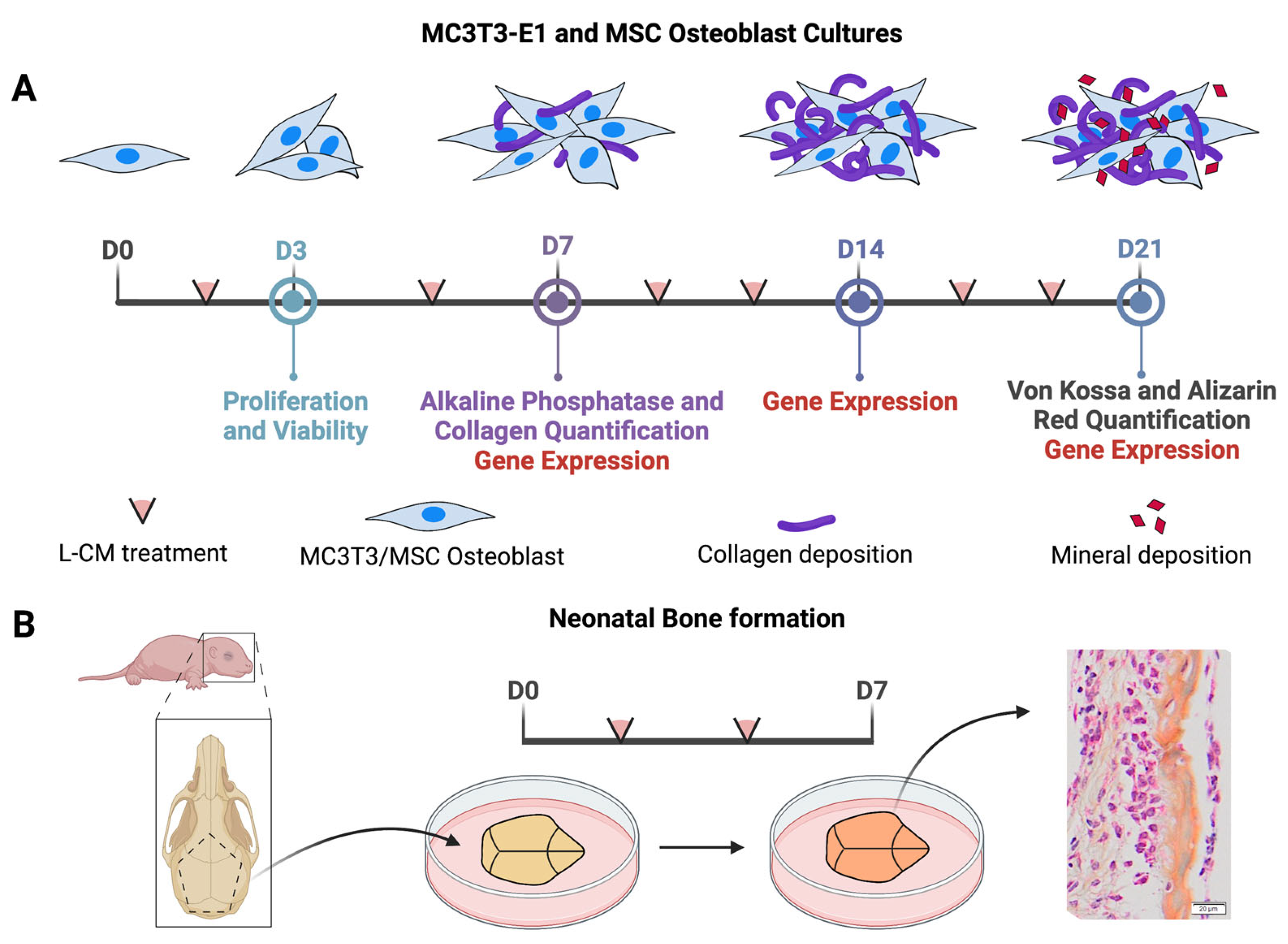
2. Materials and Methods
2.1. Mice
2.2. Lymphatic Conditioned Medium (L-CM) Preparation
2.3. Proliferation and Viability
2.4. Isolation of Primary Osteoblasts and Differentiation
2.5. Alkaline Phosphatase (ALP) and Collagen Staining and Quantification
2.6. Von Kossa and Alizarin Red Staining and Quantification
2.7. RNA Isolation and RT-qPCR Analysis
2.8. Organ Culture, Staining, and Quantification
2.9. Statistical Analysis
3. Results
3.1. L-CM Treatment Decreased MC3T3-E1 Cell Proliferation
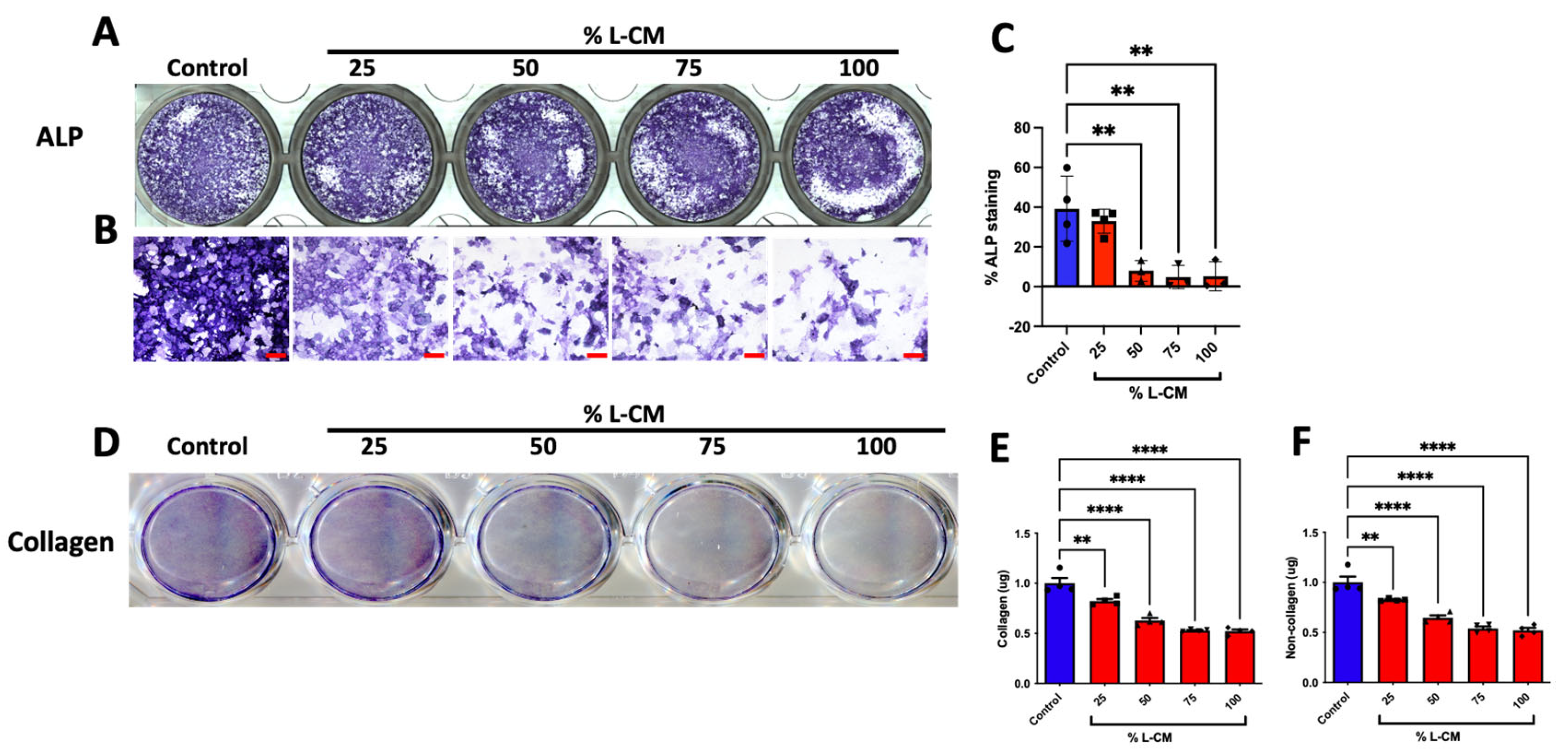
3.2. L-CM Treatment Inhibited Early Differentiation and Collagen Production in MC3T3-E1
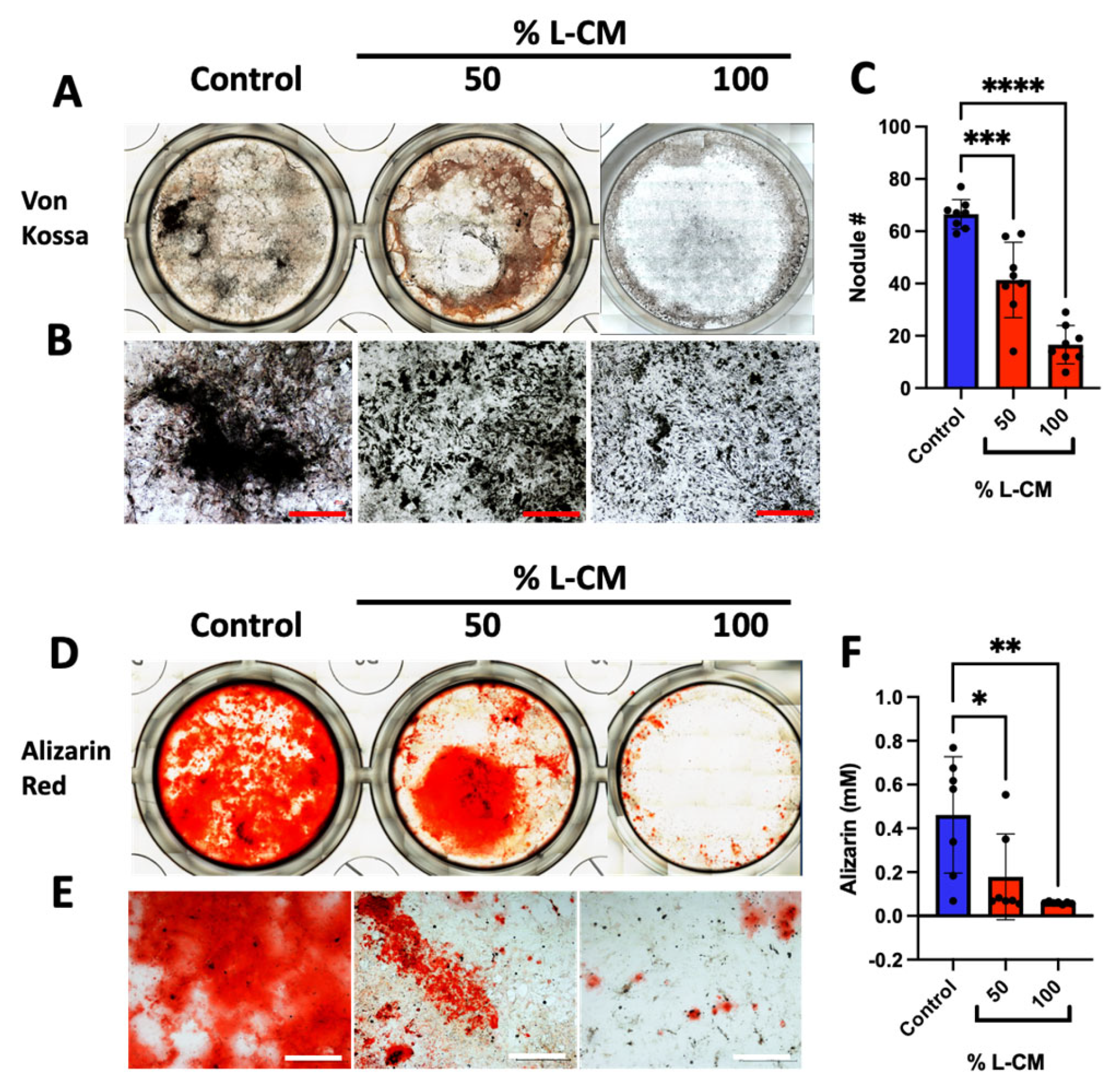
3.3. L-CM Treatment Inhibited MC3T3-E1 Matrix Mineral Deposition
3.4. L-CM Treatment Decreased the Gene Expression of Osteoblast-Related Markers
3.5. L-CM Treatment Inhibited Bone-Marrow-Derived MSC Proliferation
3.6. L-CM Treatment Decreased Early Differentiation and Collagen Production of Bone-Marrow-Derived Osteoblasts
3.7. L-CM Treatment Decreased Bone-Marrow-Derived Osteoblast Matrix Mineral Deposition
3.8. Co-Culture with Lymphatic Endothelial Cells Inhibited Osteoblast Differentiation
3.9. L-CM Inhibited Bone Formation in Calvaria Organ Culture
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Solorzano, E.; Alejo, A.L.; Ball, H.C.; Magoline, J.; Khalil, Y.; Kelly, M.; Safadi, F.F. Osteopathy in Complex Lymphatic Anomalies. Int. J. Mol. Sci. 2022, 23, 8258. [Google Scholar] [CrossRef] [PubMed]
- Heffez, L.; Doku, H.C.; Carter, B.L.; Feeney, J.E. Perspectives on massive osteolysis. Report of a case and review of the literature. Oral. Surg. Oral. Med. Oral. Pathol. 1983, 55, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Saify, F.Y.; Gosavi, S.R. Gorham’s disease: A diagnostic challenge. J. Oral. Maxillofac. Pathol. 2014, 18, 411–414. [Google Scholar] [CrossRef] [PubMed]
- Nikolaou, V.S.; Chytas, D.; Korres, D.; Efstathopoulos, N. Vanishing bone disease (Gorham-Stout syndrome): A review of a rare entity. World J. Orthop. 2014, 5, 694–698. [Google Scholar] [CrossRef] [PubMed]
- Angelini, A.; Mosele, N.; Pagliarini, E.; Ruggieri, P. Current concepts from diagnosis to management in Gorham-Stout disease: A systematic narrative review of about 350 cases. EFORT Open Rev. 2022, 7, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Homayun-Sepehr, N.; McCarter, A.L.; Helaers, R.; Galant, C.; Boon, L.M.; Brouillard, P.; Vikkula, M.; Dellinger, M.T. KRAS-driven model of Gorham-Stout disease effectively treated with trametinib. JCI Insight 2021, 6, 149831. [Google Scholar] [CrossRef] [PubMed]
- Manevitz-Mendelson, E.; Leichner, G.S.; Barel, O.; Davidi-Avrahami, I.; Ziv-Strasser, L.; Eyal, E.; Pessach, I.; Rimon, U.; Barzilai, A.; Hirshberg, A.; et al. Somatic NRAS mutation in patient with generalized lymphatic anomaly. Angiogenesis 2018, 21, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Ozeki, M.; Fukao, T.; Matsuo, M. MR imaging findings of vertebral involvement in Gorham-Stout disease, generalized lymphatic anomaly, and kaposiform lymphangiomatosis. Jpn. J. Radiol. 2017, 35, 606–612. [Google Scholar] [CrossRef]
- Trenor, C.C., 3rd; Chaudry, G. Complex lymphatic anomalies. Semin. Pediatr. Surg. 2014, 23, 186–190. [Google Scholar] [CrossRef]
- Alderfer, L.; Wei, A.; Hanjaya-Putra, D. Lymphatic Tissue Engineering and Regeneration. J. Biol. Eng. 2018, 12, 32. [Google Scholar] [CrossRef]
- Azzali, G. Transendothelial transport of lipids in the absorbing lymphatic vessel. Experientia 1982, 38, 275–277. [Google Scholar] [CrossRef] [PubMed]
- Oliver, G.; Harvey, N. A stepwise model of the development of lymphatic vasculature. Ann. N. Y. Acad. Sci. 2002, 979, 159–165; discussion 188–196. [Google Scholar] [CrossRef] [PubMed]
- Biswas, L.; Chen, J.; De Angelis, J.; Singh, A.; Owen-Woods, C.; Ding, Z.; Pujol, J.M.; Kumar, N.; Zeng, F.; Ramasamy, S.K.; et al. Lymphatic vessels in bone support regeneration after injury. Cell 2023, 186, 382–397.e324. [Google Scholar] [CrossRef] [PubMed]
- Afshari, A.; Shamdani, S.; Uzan, G.; Naserian, S.; Azarpira, N. Different approaches for transformation of mesenchymal stem cells into hepatocyte-like cells. Stem Cell Res. Ther. 2020, 11, 54. [Google Scholar] [CrossRef] [PubMed]
- Chitteti, B.R.; Cheng, Y.H.; Streicher, D.A.; Rodriguez-Rodriguez, S.; Carlesso, N.; Srour, E.F.; Kacena, M.A. Osteoblast lineage cells expressing high levels of Runx2 enhance hematopoietic progenitor cell proliferation and function. J. Cell Biochem. 2010, 111, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Amarasekara, D.S.; Kim, S.; Rho, J. Regulation of Osteoblast Differentiation by Cytokine Networks. Int. J. Mol. Sci. 2021, 22, 2851. [Google Scholar] [CrossRef] [PubMed]
- McDonald, M.M.; Khoo, W.H.; Ng, P.Y.; Xiao, Y.; Zamerli, J.; Thatcher, P.; Kyaw, W.; Pathmanandavel, K.; Grootveld, A.K.; Moran, I.; et al. Osteoclasts recycle via osteomorphs during RANKL-stimulated bone resorption. Cell 2021, 184, 1330–1347.e1313. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.H. Osteoblast-Osteoclast Communication and Bone Homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef]
- Hirayama, T.; Sabokbar, A.; Itonaga, I.; Watt-Smith, S.; Athanasou, N.A. Cellular and humoral mechanisms of osteoclast formation and bone resorption in Gorham–Stout disease. J. Pathol. 2001, 195, 624–630. [Google Scholar] [CrossRef]
- Möller, G.; Priemel, M.; Amling, M.; Werner, M.; Kuhlmey, A.S.; Delling, G. The Gorham-Stout syndrome (Gorham’s massive osteolysis). J. Bone Jt. Surg. Br. Vol. 1999, 81-B, 501–506. [Google Scholar] [CrossRef]
- Edwards, J.R.; Williams, K.; Kindblom, L.G.; Meis-Kindblom, J.M.; Hogendoorn, P.C.; Hughes, D.; Forsyth, R.G.; Jackson, D.; Athanasou, N.A. Lymphatics and bone. Hum. Pathol. 2008, 39, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wu, Y.; Liang, Y.; Tan, W.; Liu, Z.; Xiao, J. Regulation of expression level of fms-like tyrosine kinase-4 is related to osteoclast differentiation. Arch. Med. Sci. 2016, 12, 502–506. [Google Scholar] [CrossRef]
- Jones, D.; Min, W. An overview of lymphatic vessels and their emerging role in cardiovascular disease. J. Cardiovasc. Dis. Res. 2011, 2, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Monroy, M.; McCarter, A.L.; Hominick, D.; Cassidy, N.; Dellinger, M.T. Lymphatics in bone arise from pre-existing lymphatics. Development 2020, 147, dev184291. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.; Schulze, E.; Sabokbar, A.; Gordon-Andrews, H.; Jackson, D.; Athanasou, N.A. Absence of lymphatics at the bone-implant interface: Implications for periprosthetic osteolysis. Acta Orthop. 2008, 79, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, H.; Zhou, X.; Li, X.; Sun, W.; Dellinger, M.; Boyce, B.F.; Xing, L. Lymphatic Endothelial Cells Produce M-CSF, Causing Massive Bone Loss in Mice. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2019, 34, 2162. [Google Scholar] [CrossRef] [PubMed]
- Devlin, R.D.; Bone, H.G., 3rd; Roodman, G.D. Interleukin-6: A potential mediator of the massive osteolysis in patients with Gorham-Stout disease. J. Clin. Endocrinol. Metab. 1996, 81, 1893–1897. [Google Scholar] [PubMed]
- Rossi, M.; Buonuomo, P.S.; Battafarano, G.; Conforti, A.; Mariani, E.; Algeri, M.; Pelle, S.; D’Agostini, M.; Macchiaiolo, M.; De Vito, R.; et al. Dissecting the mechanisms of bone loss in Gorham-Stout disease. Bone 2020, 130, 115068. [Google Scholar] [CrossRef]
- Brodszki, N.; Länsberg, J.K.; Dictor, M.; Gyllstedt, E.; Ewers, S.B.; Larsson, M.K.; Eklund, E.A. A novel treatment approach for paediatric Gorham-Stout syndrome with chylothorax. Acta Paediatr. 2011, 100, 1448–1453. [Google Scholar] [CrossRef]
- Abdelmagid, S.M.; Belcher, J.Y.; Moussa, F.M.; Lababidi, S.L.; Sondag, G.R.; Novak, K.M.; Sanyurah, A.S.; Frara, N.A.; Razmpour, R.; Del Carpio-Cano, F.E.; et al. Mutation in osteoactivin decreases bone formation in vivo and osteoblast differentiation in vitro. Am. J. Pathol. 2014, 184, 697–713. [Google Scholar] [CrossRef]
- Ball, H.C.; Moussa, F.M.; Mbimba, T.; Orman, R.; Safadi, F.F.; Cooper, L.N. Methods and insights from the characterization of osteoprogenitor cells of bats (Mammalia: Chiroptera). Stem Cell Res. 2016, 17, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Moussa, F.M.; Cook, B.P.; Sondag, G.R.; DeSanto, M.; Obri, M.S.; McDermott, S.E.; Safadi, F.F. The role of miR-150 regulates bone cell differentiation and function. Bone 2021, 145, 115470. [Google Scholar] [CrossRef] [PubMed]
- Abdelmagid, S.M.; Barbe, M.F.; Rico, M.C.; Salihoglu, S.; Arango-Hisijara, I.; Selim, A.H.; Anderson, M.G.; Owen, T.A.; Popoff, S.N.; Safadi, F.F. Osteoactivin, an anabolic factor that regulates osteoblast differentiation and function. Exp. Cell Res. 2008, 314, 2334–2351. [Google Scholar] [CrossRef] [PubMed]
- Safadi, F.F.; Xu, J.; Smock, S.L.; Kanaan, R.A.; Selim, A.H.; Odgren, P.R.; Marks, S.C., Jr.; Owen, T.A.; Popoff, S.N. Expression of connective tissue growth factor in bone: Its role in osteoblast proliferation and differentiation in vitro and bone formation in vivo. J. Cell. Physiol. 2003, 196, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Sondag, G.R.; Mbimba, T.S.; Moussa, F.M.; Novak, K.; Yu, B.; Jaber, F.A.; Abdelmagid, S.M.; Geldenhuys, W.J.; Safadi, F.F. Osteoactivin inhibition of osteoclastogenesis is mediated through CD44-ERK signaling. Exp. Mol. Med. 2016, 48, e257. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Lu, Y.; Wang, Y.; Ge, L.; Zhai, N.; Han, J. A protocol for isolation and identification and comparative characterization of primary osteoblasts from mouse and rat calvaria. Cell Tissue Bank. 2019, 20, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, K.S.; Chirgwin, J.M.; Guise, T.A. Assessing New Bone Formation in Neonatal Calvarial Organ Cultures. In Osteoporosis: Methods and Protocols; Westendorf, J.J., Ed.; Humana Press: Totowa, NJ, USA, 2008; pp. 37–50. [Google Scholar]
- Alborzi, A.; Mac, K.; Glackin, C.A.; Murray, S.S.; Zernik, J.H. Endochondral and intramembranous fetal bone development: Osteoblastic cell proliferation, and expression of alkaline phosphatase, m-twist, and histone H4. J. Craniofacial Genet. Dev. Biol. 1996, 16, 94–106. [Google Scholar]
- Morais, S.; Carvalho, G.S.; Faria, J.L.; Gomes, H.T.; Sousa, J.P. In vitro biomineralization by osteoblast-like cells. II. Characterization of cellular culture supernatants. Biomaterials 1998, 19, 23–29. [Google Scholar] [CrossRef]
- Lynch, M.P.; Stein, J.L.; Stein, G.S.; Lian, J.B. The influence of type I collagen on the development and maintenance of the osteoblast phenotype in primary and passaged rat calvarial osteoblasts: Modification of expression of genes supporting cell growth, adhesion, and extracellular matrix mineralization. Exp. Cell Res. 1995, 216, 35–45. [Google Scholar] [CrossRef]
- Bills, C.E.; Eisenberg, H.; Pallante, S.L. Complexes of organic acids with calcium phosphate: The Von Kossa stain as a clue to the composition of bone mineral. Johns Hopkins Med. J. 1974, 128, 194–207. [Google Scholar]
- Driessens, F.C.; van Dijk, J.W.; Borggreven, J.M. Biological calcium phosphates and their role in the physiology of bone and dental tissues I. Composition and solubility of calcium phosphates. Calcif. Tissue Res. 1978, 26, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Prince, M.; Banerjee, C.; Javed, A.; Green, J.; Lian, J.B.; Stein, G.S.; Bodine, P.V.; Komm, B.S. Expression and regulation of Runx2/Cbfa1 and osteoblast phenotypic markers during the growth and differentiation of human osteoblasts. J. Cell Biochem. 2001, 80, 424–440. [Google Scholar] [CrossRef] [PubMed]
- Bateman, J.P.; Safadi, F.F.; Susin, C.; Wikesjo, U.M. Exploratory study on the effect of osteoactivin on bone formation in the rat critical-size calvarial defect model. J. Periodontal Res. 2012, 47, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Frara, N.; Abdelmagid, S.M.; Sondag, G.R.; Moussa, F.M.; Yingling, V.R.; Owen, T.A.; Popoff, S.N.; Barbe, M.F.; Safadi, F.F. Transgenic Expression of Osteoactivin/gpnmb Enhances Bone Formation In Vivo and Osteoprogenitor Differentiation Ex Vivo. J. Cell. Physiol. 2016, 231, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Abdelmagid, S.M.; Sondag, G.R.; Moussa, F.M.; Belcher, J.Y.; Yu, B.; Stinnett, H.; Novak, K.; Mbimba, T.; Khol, M.; Hankenson, K.D.; et al. Mutation in Osteoactivin Promotes Receptor Activator of NFkappaB Ligand (RANKL)-mediated Osteoclast Differentiation and Survival but Inhibits Osteoclast Function. J. Biol. Chem. 2015, 290, 20128–20146. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Udagawa, N.; Suda, T. A new member of tumor necrosis factor ligand family, ODF/OPGL/TRANCE/RANKL, regulates osteoclast differentiation and function. Biochem. Biophys. Res. Commun. 1999, 256, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Gori, F.; Hofbauer, L.C.; Dunstan, C.R.; Spelsberg, T.C.; Khosla, S.; Riggs, B.L. The expression of osteoprotegerin and RANK ligand and the support of osteoclast formation by stromal-osteoblast lineage cells is developmentally regulated. Endocrinology 2000, 141, 4768–4776. [Google Scholar] [CrossRef]
- Schneider, K.N.; Masthoff, M.; Gosheger, G.; Klingebiel, S.; Schorn, D.; Roder, J.; Vogler, T.; Wildgruber, M.; Andreou, D. Gorham-Stout disease: Good results of bisphosphonate treatment in 6 of 7 patients. Acta Orthop. 2020, 91, 209–214. [Google Scholar] [CrossRef]
- Arosarena, O.A.; Barr, E.W.; Thorpe, R.; Yankey, H.; Tarr, J.T.; Safadi, F.F. Osteoactivin regulates head and neck squamous cell carcinoma invasion by modulating matrix metalloproteases. J. Cell. Physiol. 2018, 233, 409–421. [Google Scholar] [CrossRef]
- Yu, B.; Sondag, G.R.; Malcuit, C.; Kim, M.H.; Safadi, F.F. Macrophage-Associated Osteoactivin/GPNMB Mediates Mesenchymal Stem Cell Survival, Proliferation, and Migration Via a CD44-Dependent Mechanism. J. Cell Biochem. 2016, 117, 1511–1521. [Google Scholar] [CrossRef]
- Blair, H.C.; Larrouture, Q.C.; Li, Y.; Lin, H.; Beer-Stoltz, D.; Liu, L.; Tuan, R.S.; Robinson, L.J.; Schlesinger, P.H.; Nelson, D.J. Osteoblast Differentiation and Bone Matrix Formation In Vivo and In Vitro. Tissue Eng. Part. B Rev. 2017, 23, 268–280. [Google Scholar] [CrossRef]
- Karner, C.M.; Long, F.X. Wnt signaling and cellular metabolism in osteoblasts. Cell Mol. Life Sci. 2017, 74, 1649–1657. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Park, S.; Roh, S. Transdifferentiation of mouse adipose-derived stromal cells into acinar cells of the submandibular gland using a co-culture system. Exp. Cell Res. 2015, 334, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Nicolin, V.; Bortul, R.; Bareggi, R.; Baldini, G.; Martinelli, B.; Narducci, P. Breast adenocarcinoma MCF-7 cell line induces spontaneous osteoclastogenesis via a RANK-ligand-dependent pathway. Acta Histochem. 2008, 110, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Harris-Hooker, S.; Kumar, R.; Sanford, G. Co-culture of Retinal and Endothelial Cells Results in the Modulation of Genes Critical to Retinal Neovascularization. Vasc. Cell 2011, 3, 27. [Google Scholar] [CrossRef] [PubMed]
- Gem, M.; Ozkul, E.; Arslan, H. Gorham-Stout’s disease in the metatarsus: A case report. Acta Orthop. Traumatol. Turc. 2014, 48, 467–471. [Google Scholar] [CrossRef] [PubMed]
- Tena-Sanabria, M.E.; Jesús-Mejenes, L.Y.; Fuentes-Herrera, G.; Álvarez-Martínez, F.A.; Victorio-García, N.P.; Núñez-Enríquez, J.C. A report of two children with Gorham-Stout disease. BMC Pediatr. 2019, 19, 206. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Rana, I.; Buonuomo, P.S.; Battafarano, G.; Mariani, E.; D’Agostini, M.; Porzio, O.; De Martino, V.; Minisola, S.; Macchiaiolo, M.; et al. Dysregulated miRNAs in bone cells of patients with Gorham-Stout disease. FASEB J. 2021, 35, e21424. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; March, M.E.; Gutierrez-Uzquiza, A.; Kao, C.; Seiler, C.; Pinto, E.; Matsuoka, L.S.; Battig, M.R.; Bhoj, E.J.; Wenger, T.L.; et al. ARAF recurrent mutation causes central conducting lymphatic anomaly treatable with a MEK inhibitor. Nat. Med. 2019, 25, 1116–1122. [Google Scholar] [CrossRef]
- Rodriguez-Laguna, L.; Agra, N.; Ibanez, K.; Oliva-Molina, G.; Gordo, G.; Khurana, N.; Hominick, D.; Beato, M.; Colmenero, I.; Herranz, G.; et al. Somatic activating mutations in PIK3CA cause generalized lymphatic anomaly. J. Exp. Med. 2019, 216, 407–418. [Google Scholar] [CrossRef]
- Ozeki, M.; Aoki, Y.; Nozawa, A.; Yasue, S.; Endo, S.; Hori, Y.; Matsuoka, K.; Niihori, T.; Funayama, R.; Shirota, M.; et al. Detection of NRAS mutation in cell-free DNA biological fluids from patients with kaposiform lymphangiomatosis. Orphanet J. Rare Dis. 2019, 14, 215. [Google Scholar] [CrossRef] [PubMed]
- Cueni, L.N.; Detmar, M. New insights into the molecular control of the lymphatic vascular system and its role in disease. J. Investig. Dermatol. 2006, 126, 2167–2177. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Oliver, G. Current views on the function of the lymphatic vasculature in health and disease. Genes. Dev. 2010, 24, 2115–2126. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, A.; Detmar, M. Lymphangiogenesis and cancer. Genes. Cancer 2011, 2, 1146–1158. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Kirk, M.; Kahn, A.J. The role of type I collagen in the regulation of the osteoblast phenotype. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 1996, 11, 1139–1145. [Google Scholar] [CrossRef] [PubMed]
- Aubin, J.E.; Liu, F.; Malaval, L.; Gupta, A.K. Osteoblast and chondroblast differentiation. Bone 1995, 17 (Suppl. S2), 77s–83s. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, M.; Fujisawa, R.; Kuboki, Y. Type I collagen-induced osteoblastic differentiation of bone-marrow cells mediated by collagen-alpha2beta1 integrin interaction. J. Cell. Physiol. 2000, 184, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Maehata, Y.; Lee, M.-C.-i.; Hata, R.-I. Roles of Collagen Molecules in Growth and Differentiation of Human Osteoblasts. J. Oral. Biosci. 2009, 51, 72–80. [Google Scholar] [CrossRef]
- Patntirapong, S.; Chanruangvanit, C.; Lavanrattanakul, K.; Satravaha, Y. Assessment of bisphosphonate treated-osteoblast behaviors by conventional assays and a simple digital image analysis. Acta Histochem. 2021, 123, 151659. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequence |
|---|---|
| m_18s_F | CTTAGAGGGACAAGTGGCG |
| m_18s_R | ACGCTGAGCCAGTCAGTGTA |
| m_ALP_F | CCAACTCTTTTGTGCCAGAGA |
| m_ALP_R | GGCTACATTGGTGTTGAGCTTTT |
| m_Col1 A1_F | GCTCCTCTTAGGGGCCACT |
| m_Col1 A1_R | ATTGGGGACCCTTAGGCCAT |
| m_GPNMB_F | AATGGGTCTGGCACCTACTG |
| m_GPNMB_R | GGCTTGTACGCCTTGTGTTT |
| m_OPG_R | AGCAGGAGTGCAACCGCACC |
| m_OPG_F | TTCCAGCTTGCACCACGCCG |
| m_RANK-L_F | GCTCCGAGCTGGTGAAGAAA |
| m_RANK-L_R | CCCCAAAGTACGTCGCATCT |
| m_Runx2_F | GACTGTGGTTACCGTCATGGC |
| m_Runx2_R | ACTTGGTTTTTCATAACAGCGGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solorzano, E.; Alejo, A.L.; Ball, H.C.; Robinson, G.T.; Solorzano, A.L.; Safadi, R.; Douglas, J.; Kelly, M.; Safadi, F.F. The Lymphatic Endothelial Cell Secretome Inhibits Osteoblast Differentiation and Bone Formation. Cells 2023, 12, 2482. https://doi.org/10.3390/cells12202482
Solorzano E, Alejo AL, Ball HC, Robinson GT, Solorzano AL, Safadi R, Douglas J, Kelly M, Safadi FF. The Lymphatic Endothelial Cell Secretome Inhibits Osteoblast Differentiation and Bone Formation. Cells. 2023; 12(20):2482. https://doi.org/10.3390/cells12202482
Chicago/Turabian StyleSolorzano, Ernesto, Andrew L. Alejo, Hope C. Ball, Gabrielle T. Robinson, Andrea L. Solorzano, Rama Safadi, Jacob Douglas, Michael Kelly, and Fayez F. Safadi. 2023. "The Lymphatic Endothelial Cell Secretome Inhibits Osteoblast Differentiation and Bone Formation" Cells 12, no. 20: 2482. https://doi.org/10.3390/cells12202482