
Sanguisorba minor Scop.: An Overview of Its Phytochemistry and Biological Effects
 , ,
, ,
Abstract
:1. Introduction
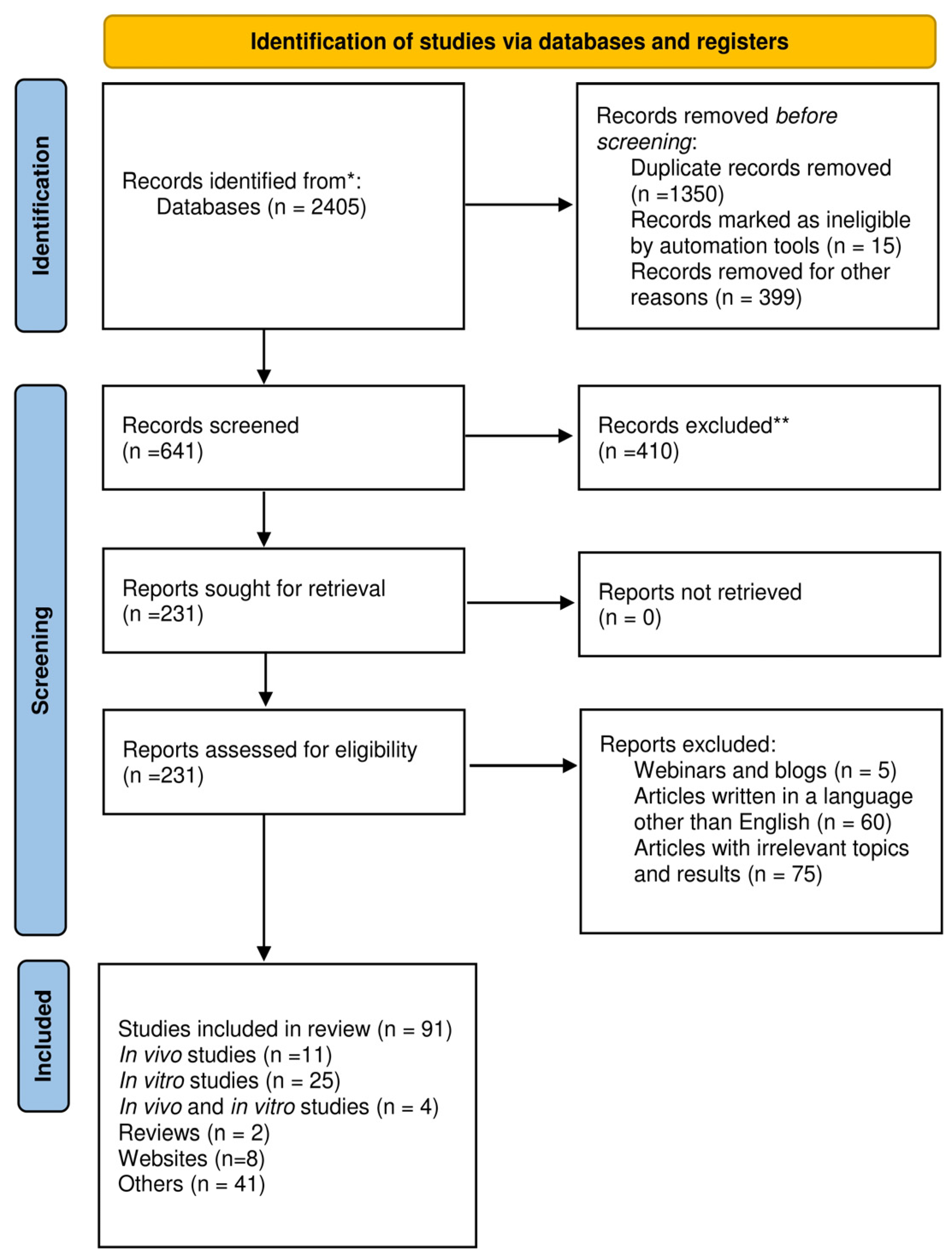
Research Methodology
2. Taxonomy, Habitat, and Distribution
3. Botanical Characterization: Macroscopic and Microscopic Aspects of S. minor

4. The Nutritional Composition of S. minor
5. Phytochemical Composition of Botanical Part of S. minor
6. Antioxidant Capacity
7. Biological Activities
7.1. Antimicrobial Effects
7.1.1. Antifungal Activity
7.1.2. Antiviral Activity
7.2. Cytotoxic Activity on Cancer Cells
7.3. Neuroprotective Effects
7.4. Antiulcerogenic Activity
7.5. Toxicity Studies
8. Insects: Pest or Beneficial
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, P.; Li, J.; Chen, Q.; Wang, L.; Yang, J.; Wu, A.; Jiang, N.; Liu, Y.; Chen, J.; Zou, W.; et al. A Comprehensive Review of Genus Sanguisorba: Traditional Uses, Chemical Constituents and Medical Applications. Front. Pharmacol. 2021, 12, 750165. [Google Scholar] [CrossRef] [PubMed]
- The Angiosperm Phylogeny Group. An Update of the Angiosperm Phylogeny Group Classification for the Orders and Families of Flowering Plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef]
- Wang, J.-R.; Wang, X.; Su, N.; Li, Q.-J.; Zhang, X.-H.; Ma, Y.-P.; Zhao, L.; Ginefra Toni, J.F.; De Craene, L.R. Floral Morphology and Morphogenesis in Sanguisorba (Rosaceae): Flower Diversification despite Petal Reduction and Spatial Constraints. Bot. J. Linn. Soc. 2020, 193, 47–63. [Google Scholar] [CrossRef]
- Tocai, A.C.; Memete, A.R.; Vicaş, S.; Burescu, P. Antioxidant Capacity of Sanguisorba officinalis L. and Sanguisorba Minor Scop. NRSD 2021, 11, 121–133. [Google Scholar] [CrossRef]
- Zhang, F.; Fu, T.-J.; Peng, S.-L.; Liu, Z.-R.; Ding, L.-S. Two New Triterpenoids from the Roots of Sanguisorba officinalis L. J. Integr. Plant Biol. 2005, 47, 251–256. [Google Scholar] [CrossRef]
- Paniagua-Zambrana, N.Y.; Bussmann, R.W.; Romero, C. Sanguisorba Minor Scop. Rosaceae. In Ethnobotany of the Andes; Paniagua-Zambrana, N.Y., Bussmann, R.W., Eds.; Ethnobotany of Mountain Regions; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–5. ISBN 978-3-319-77093-2. [Google Scholar]
- Ciubotaru, A. National Botanical Garden (Institute); Tofan-Dorofeev, E. Genurile sanguisorba L. Și poterium L. (Rosaceae Adans.) În Republica Moldova. J. Bot. 2021, 21, 155–156. [Google Scholar] [CrossRef]
- Guarrera, P.M.; Savo, V. Wild Food Plants Used in Traditional Vegetable Mixtures in Italy. J. Ethnopharmacol. 2016, 185, 202–234. [Google Scholar] [CrossRef]
- Zhang, Y.; MacAdam, J.W.; Villalba, J.J.; Dai, X. In Vitro Digestibility of Mountain-grown Irrigated Perennial Legume, Grass and Forb Forages Is Influenced by Elevated Non-fibrous Carbohydrates and Plant Secondary Compounds. J. Sci. Food Agric. 2021, 101, 334–340. [Google Scholar] [CrossRef]
- Scopoli, G.A.; Krauss, J.P.; Rein, J.F. Ioannis Antonii Scopoli... Flora Carniolica: Exhibens Plantas Carnioliae Indigenas et Distributas in Classes, Genera, Species, Varietates, Ordine Linnaeano; Impensis Ioannis Pauli Krauss, Bibliopolae Vindobonensis: Vienna, Austria, 1772. [Google Scholar]
- Cirovic, T.; Barjaktarevic, A.; Cupara, S.; Mitic, V.; Nikolic, J.; Jovanovic, V.S. Antioxidant and Antimicrobial Activity of Sanguisorba Minor L. Extracts. Serb. J. Exp. Clin. Res. 2022, 23, 51–57. [Google Scholar] [CrossRef]
- Gerard, J. The Herball: Or, Generall Historie of Plants; The English experience, its record in early printed books published in facsimile; Theatrum Orbis Terrarum; W. J. Johnson: Amsterdam, The Netherlands; Norwood, NJ, USA, 1974; ISBN 978-90-221-9998-5. [Google Scholar]
- Traian, S. Flora României; Edit; Academiei Române: Bucharest, Romania, 1956; Volume 4. [Google Scholar]
- Viano, J.; Masotti, V.; Gaydou, E.M. Nutritional Value of Mediterranean Sheep’s Burnet (Sanguisorba Minor Ssp. Muricata ). J. Agric. Food Chem. 1999, 47, 4645–4648. [Google Scholar] [CrossRef]
- Zhao, Z.; He, X.; Zhang, Q.; Wei, X.; Huang, L.; Fang, J.C.; Wang, X.; Zhao, M.; Bai, Y.; Zheng, X. Traditional Uses, Chemical Constituents and Biological Activities of Plants from the Genus Sanguisorba L. Am. J. Chin. Med. 2017, 45, 199–224. [Google Scholar] [CrossRef] [PubMed]
- Sabbatini, A.; Jurnatan, Y.; Fraatz, M.A.; Govori, S.; Haziri, A.; Millaku, F.; Zorn, H.; Zhang, Y. Aroma Characterization of a Wild Plant (Sanguisorba Albanica) from Kosovo Using Multiple Headspace Solid Phase Microextraction Combined with Gas Chromatography-Mass Spectrometry-Olfactometry. Food Res. Int. 2019, 120, 514–522. [Google Scholar] [CrossRef]
- Nordborg, G. Sanguisorba L., Sarcopoterium Spach and Bencomia Webb et Berth: Delimitation and Subdivision of the Genera; Distributor, Almqvist & Wiksell: Stockholm, Sweden, 1966; Volume 2. [Google Scholar]
- Ardelean, A.; Mohan, G. Flora medicinală a României; Editura ALL: Bucureşti, Romania, 2008; ISBN 978-973-571-838-1. [Google Scholar]
- Karkanis, A.; Vellios, E.; Thomaidis, T.; Bilalis, D.; Efthimiadou, A.; Travlos, I. Phytochemistry and Biological Properties of Burnet Weed (Sanguisorba Spp.): A Review. Not. Sci. Biol. 2014, 6, 395–398. [Google Scholar] [CrossRef]
- Gerard, J. The Herbal or General History of Plants: The Complete 1633 Edition as Revised and Enlarged by Thomas Johnson, Revised-Enlarged ed.; Dover Publications: Mineola, New York, NY, USA, 2015; ISBN 978-1-60660-080-1. [Google Scholar]
- Guarrera, P.M.; Savo, V. Perceived Health Properties of Wild and Cultivated Food Plants in Local and Popular Traditions of Italy: A Review. J. Ethnopharmacol. 2013, 146, 659–680. [Google Scholar] [CrossRef] [PubMed]
- Fierascu, R.C.; Fierascu, I.; Ortan, A.; Avramescu, S.M.; Dinu-Pirvu, C.E.; Ionescu, D. Romanian Aromatic and Medicinal Plants: From Tradition to Science. In Aromatic and Medicinal Plants—Back to Nature; El-Shemy, H.A., Ed.; InTech: Bucharest, Romania, 2017; ISBN 978-953-51-2977-6. [Google Scholar]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Bunse, M.; Stintzing, F.; Kammerer, D.R. Morphology and Phytochemistry of Sanguisorba Officinalis L. Seeds (Rosaceae). J. Appl. Bot. Food Qual. 2021, 94, 92–98. [Google Scholar] [CrossRef]
- Cirovic, T.; Barjaktarevic, A.; Ninkovic, M.; Bauer, R.; Nikles, S.; Brankovic, S.; Markovic, M.; Stankov Jovanovic, V.; Ilic, M.; Milovanovic, O.; et al. Biological Activities of Sanguisorba minor L. Extracts—In vitro and In vivo Evaluations. Acta Pol. Pharm. Drug Res. 2020, 77, 745–758. [Google Scholar] [CrossRef]
- Mishima, M.; Ohmido, N.; Fukui, K.; Yahara, T. Trends in Site-Number Change of RDNA Loci during Polyploid Evolution in Sanguisorba (Rosaceae). Chromosoma 2002, 110, 550–558. [Google Scholar] [CrossRef]
- Lee, S.; Heo, K.; Cho, J.; Lee, C.; Chen, W.; Kim, S.-C. New Insights into Pollen Morphology and Its Implications in the Phylogeny of Sanguisorba L. (Rosaceae; Sanguisorbeae). Plant. Syst. Evol. 2011, 291, 227–242. [Google Scholar] [CrossRef]
- Karkanis, A.C.; Fernandes, Â.; Vaz, J.; Petropoulos, S.; Georgiou, E.; Ciric, A.; Sokovic, M.; Oludemi, T.; Barros, L.; Ferreira, I.C.F.R. Chemical Composition and Bioactive Properties of Sanguisorba Minor Scop. under Mediterranean Growing Conditions. Food Funct. 2019, 10, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Ceccanti, C.; Landi, M.; Rocchetti, G.; Miras Moreno, M.B.; Lucini, L.; Incrocci, L.; Pardossi, A.; Guidi, L. Hydroponically Grown Sanguisorba Minor Scop.: Effects of Cut and Storage on Fresh-Cut Produce. Antioxidants 2019, 8, 631. [Google Scholar] [CrossRef] [PubMed]
- Singhurst, J.R.; Holmes, W.C. Sanguisorba Minor (Rosaceae) Adventive in Texas. Phytoneuron 2012, 93, 1–3. [Google Scholar]
- Calone, R.; Bregaglio, S.; Sanoubar, R.; Noli, E.; Lambertini, C.; Barbanti, L. Physiological Adaptation to Water Salinity in Six Wild Halophytes Suitable for Mediterranean Agriculture. Plants 2021, 10, 309. [Google Scholar] [CrossRef] [PubMed]
- Çöçü, S.; Khawar, K.M.; Sancak, C.; Özcan, S.; Bornman, C.H.; Gülbitti-Onarici, S. Plant Regeneration in Vitro from Immature Embryos of Lesser Burnet (Sanguisorba Minor Scop.). South. Afr. J. Bot. 2003, 69, 446–447. [Google Scholar] [CrossRef]
- Farmahini Farahani, A.; Tavili, A.; Azarnivand, H.; Jafari, A. Effect of Priming and Nano Particles Application on Seedling Emergence, Establishment, Growth and Physiological Characteristics of Sanguisorba Minor Scop. and Agropyron Intermedium (Host) P. Beauv Forage Species under Drought Stress in Natural Field. Rangeland 2022, 16, 236–250. [Google Scholar]
- Torres, C.D.; Magnin, A.; Sabatier, S.; Puntieri, J.G.; Caraglio, Y. Assessing Coordinated Intra-Specific Variation in Root/Shoot Traits in Two Herbaceous Species Based on Architecture and Ontogeny. Folia Geobot. 2022, 57, 167–180. [Google Scholar] [CrossRef]
- World Flora Online Consortium. Available online: http://www.worldfloraonline.org/organisation/WFO (accessed on 20 April 2023).
- Sanguisorba Minor Subsp. Balearica (Nyman) Muñoz Garm. & C.Navarro. Available online: http://www.worldfloraonline.org/taxon/wfo-0001005675 (accessed on 20 April 2023).
- Sanguisorba Minor Subsp. Lasiocarpa (Boiss. & Hausskn.) Nordborg. Available online: http://www.worldfloraonline.org/taxon/wfo-0001001052 (accessed on 20 April 2023).
- Sanguisorba Minor Subsp. Magnolii (Spach) Briq. Available online: http://www.worldfloraonline.org/taxon/wfo-0001001057 (accessed on 20 April 2023).
- Sanguisorba Minor Subsp. Mauritanica (Desf.) Cout. Available online: http://www.worldfloraonline.org/taxon/wfo-0001007751 (accessed on 20 April 2023).
- Sanguisorba Minor Subsp. Muricata (Spach) Nordborg. Available online: http://www.worldfloraonline.org/taxon/wfo-0001015027 (accessed on 20 April 2023).
- Sanguisorba Minor Subsp. Verrucosa (Ehrenb. Ex Decne.) Holmboe. Available online: http://www.worldfloraonline.org/taxon/wfo-0001011229 (accessed on 20 April 2023).
- Sanguisorba Minor Scop. Available online: http://www.worldfloraonline.org/taxon/wfo-0001015888#preferredNames (accessed on 20 April 2023).
- Andrabi, S.M.; Rehman, W.; Reshi, Z.A.; Nasqhi, A.R.; Ganie, A.H. Sanguisorba Minor Scop. (Rosaceae), A New Addition to the Indian Flora. Taiwania 2012, 57, 410–412. [Google Scholar]
- Tocai (Moţoc), A.-C.; Ranga, F.; Teodorescu, A.G.; Pallag, A.; Vlad, A.M.; Bandici, L.; Vicas, S.I. Evaluation of Polyphenolic Composition and Antimicrobial Properties of Sanguisorba Officinalis L. and Sanguisorba Minor Scop. Plants 2022, 11, 3561. [Google Scholar] [CrossRef]
- Simpson, M.G. 8—Diversity and Classification of Flowering Plants: Eudicots. In Plant Systematics, 2nd ed.; Simpson, M.G., Ed.; Academic Press: San Diego, CA, USA, 2010; pp. 275–448. ISBN 978-0-12-374380-0. [Google Scholar]
- Byng, J.W. The Flowering Plants Handbook; Plant Gateway Ltd.: Bradford, UK, 2014. [Google Scholar]
- Park, I.; Song, J.; Yang, S.; Choi, G.; Moon, B. A Comprehensive Study of the Genus Sanguisorba (Rosaceae) Based on the Floral Micromorphology, Palynology, and Plastome Analysis. Genes 2021, 12, 1764. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, X.; Zhang, Z.-L.; He, L.; Wang, Z.; Wang, G.-S. Isolation and Identification of the Phenolic Compounds from the Roots of Sanguisorba Officinalis L. and Their Antioxidant Activities. Molecules 2012, 17, 13917–13922. [Google Scholar] [CrossRef]
- Cuccioloni, M.; Bonfili, L.; Mozzicafreddo, M.; Cecarini, V.; Eleuteri, A.M.; Angeletti, M. Sanguisorba Minor Extract Suppresses Plasmin-Mediated Mechanisms of Cancer Cell Migration. Biochim. Biophys. Acta Gen. Subj. 2012, 1820, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, M.; Tutar, H.; Kardes, Y.M.; Das, A.; Kokten, K. Fatty Acid Compositions of the Seeds of Different Sanguisorba Minor Genotypes. Prog. Nutr. 2019, 21, 868–870. [Google Scholar] [CrossRef]
- Holloway, P.S.; Matheke, G.E. Seed Germination of Burnet, Sanguisorba Spp. Nativ. Plants J. 2003, 4, 95–99. [Google Scholar] [CrossRef]
- Hasanović, M.; Čakar, J.; Ahatović, A.; Murtić, S.; Subašić, M.; Bajrović, K.; Durmić-Pašić, A. Physiological Parameters Indicate Remarkable Survival Mechanisms of Sanguisorba Minor Scop. on Metalliferous and Non-Metalliferous Sites. Biologia 2022, 77, 1915–1929. [Google Scholar] [CrossRef]
- Richardson, F.; Brodribb, T.J.; Jordan, G.J. Amphistomatic Leaf Surfaces Independently Regulate Gas Exchange in Response to Variations in Evaporative Demand. Tree Physiol. 2017, 37, 869–878. [Google Scholar] [CrossRef]
- Yuan, J.; Wang, X.; Zhou, H.; Li, Y.; Zhang, J.; Yu, S.; Wang, M.; Hao, M.; Zhao, Q.; Liu, L.; et al. Comparison of Sample Preparation Techniques for Inspection of Leaf Epidermises Using Light Microscopy and Scanning Electronic Microscopy. Front. Plant. Sci. 2020, 11, 133. [Google Scholar] [CrossRef]
- Finimundy, T.C.; Karkanis, A.; Fernandes, Â.; Petropoulos, S.A.; Calhelha, R.; Petrović, J.; Soković, M.; Rosa, E.; Barros, L.; Ferreira, I.C.F.R. Bioactive Properties of Sanguisorba Minor L. Cultivated in Central Greece under Different Fertilization Regimes. Food Chem. 2020, 327, 127043. [Google Scholar] [CrossRef]
- Romojaro, A.; Botella, M.Á.; Obón, C.; Pretel, M.T. Nutritional and Antioxidant Properties of Wild Edible Plants and Their Use as Potential Ingredients in the Modern Diet. Int. J. Food Sci. Nutr. 2013, 64, 944–952. [Google Scholar] [CrossRef] [PubMed]
- Romojaro, A.; Serrano, M.; Pretel, M.T. Wild Edible Plants (WEP) as Antioxidant Supplements for Oils and Juices. Acta Hortic. 2018, 1194, 1039–1044. [Google Scholar] [CrossRef]
- Reher, G.; Reznicek, G.; Baumann, A. Triterpenoids from Sarcopoterium Spinosum and Sanguisorba Minor. Planta Med. 1991, 57, 506. [Google Scholar] [CrossRef]
- Ceccanti, C.; Finimundy, T.C.; Heleno, S.A.; Pires, T.C.S.P.; Calhelha, R.C.; Guidi, L.; Ferreira, I.C.F.R.; Barros, L. Differences in the Phenolic Composition and Nutraceutical Properties of Freeze Dried and Oven-Dried Wild and Domesticated Samples of Sanguisorba Minor Scop. LWT 2021, 145, 111335. [Google Scholar] [CrossRef]
- Tong, Z.; He, W.; Fan, X.; Guo, A. Biological Function of Plant Tannin and Its Application in Animal Health. Front. Vet. Sci. 2022, 8, 803657. [Google Scholar] [CrossRef]
- Traditional Herbal Medicines: A Guide to Their Safer Use. Br. J. Clin. Pharmacol. 2008, 66, 418–420. [CrossRef]
- Gatto, M.A.; Ippolito, A.; Linsalata, V.; Cascarano, N.A.; Nigro, F.; Vanadia, S.; Di Venere, D. Activity of Extracts from Wild Edible Herbs against Postharvest Fungal Diseases of Fruit and Vegetables. Postharvest Biol. Technol. 2011, 61, 72–82. [Google Scholar] [CrossRef]
- Ayoub, N.A. Unique Phenolic Carboxylic Acids from Sanguisorba Minor. Phytochemistry 2003, 63, 433–436. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, A.; Masoudi, S.; Masnabadi, N.; Rustaiyan, A.H. Chemical Constituents of the Essential Oil of Sanguisorba Minor Scop. Leaves, from Iran. J. Med. Plants 2010, 9, 67–70. [Google Scholar]
- Ferreira, A.; Proença, C.; Serralheiro, M.L.M.; Araújo, M.E.M. The in Vitro Screening for Acetylcholinesterase Inhibition and Antioxidant Activity of Medicinal Plants from Portugal. J. Ethnopharmacol. 2006, 108, 31–37. [Google Scholar] [CrossRef]
- Li, S.; Li, S.-K.; Gan, R.-Y.; Song, F.-L.; Kuang, L.; Li, H.-B. Antioxidant Capacities and Total Phenolic Contents of Infusions from 223 Medicinal Plants. Ind. Crops Prod. 2013, 51, 289–298. [Google Scholar] [CrossRef]
- Vanzani, P.; Rossetto, M.; De Marco, V.; Sacchetti, L.E.; Paoletti, M.G.; Rigo, A. Wild Mediterranean Plants as Traditional Food: A Valuable Source of Antioxidants. J. Food Sci. 2011, 76, C46–C51. [Google Scholar] [CrossRef]
- Gürbüz, I.; Özkan, A.M.; Yesilada, E.; Kutsal, O. Anti-Ulcerogenic Activity of Some Plants Used in Folk Medicine of Pinarbasi (Kayseri, Turkey). J. Ethnopharmacol. 2005, 101, 313–318. [Google Scholar] [CrossRef]
- Borrelli, F.; Izzo, A.A. The Plant Kingdom as a Source of Anti-Ulcer Remedies. Phytother. Res. 2000, 14, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Cavalu, S.; Banica, F.; Gruian, C.; Vanea, E.; Goller, G.; Simon, V. Microscopic and Spectroscopic Investigation of Bioactive Glasses for Antibiotic Controlled Release. J. Mol. Struct. 2013, 1040, 47–52. [Google Scholar] [CrossRef]
- Hosseini, Z.; Mansouritorghabeh, F.; Kakhki, F.S.H.; Hosseini, M.; Rakhshandeh, H.; Hosseini, A.; Hasanpour, M.; Iranshahi, M.; Rajabian, A. Effect of Sanguisorba Minor on Scopolamine-Induced Memory Loss in Rat: Involvement of Oxidative Stress and Acetylcholinesterase. Metab. Brain Dis. 2022, 37, 473–488. [Google Scholar] [CrossRef]
- Arihan, O.; Özbek, H.; Özkan, A.G. Anti-Inflammatory Effects of Sanguisorba Minor Scop. Subsp. Muricata (Spach) Briq. and Cirsium Libanoticum DC. Subsp. Lycaonicum (Boiss. & Heldr.) Davis & Parris in Rat. East. J. Med. 2015, 20, 81–85. [Google Scholar]
- Ormanci, O.; Akin, I.; Sahin, F.; Yucel, O.; Simon, V.; Cavalu, S.; Goller, G. Spark Plasma Sintered Al2O3–YSZ–TiO2 Composites: Processing, Characterization and in Vivo Evaluation. Mater. Sci. Eng. C 2014, 40, 16–23. [Google Scholar] [CrossRef]
- Ravipati, A.S.; Zhang, L.; Koyyalamudi, S.R.; Jeong, S.C.; Reddy, N.; Bartlett, J.; Smith, P.T.; Shanmugam, K.; Münch, G.; Wu, M.J.; et al. Antioxidant and Anti-Inflammatory Activities of Selected Chinese Medicinal Plants and Their Relation with Antioxidant Content. BMC Complement. Altern. Med. 2012, 12, 173. [Google Scholar] [CrossRef] [PubMed]
- Granato, D.; Shahidi, F.; Wrolstad, R.; Kilmartin, P.; Melton, L.D.; Hidalgo, F.J.; Miyashita, K.; van Camp, J.; Alasalvar, C.; Ismail, A.B.; et al. Antioxidant Activity, Total Phenolics and Flavonoids Contents: Should We Ban in Vitro Screening Methods? Food Chem. 2018, 264, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Al-Jaber, H.I.; Shakya, A.K.; Elagbar, Z.A. HPLC Profiling of Selected Phenolic Acids and Flavonoids in Salvia Eigii, Salvia Hierosolymitana and Salvia Viridis Growing Wild in Jordan and Their in Vitro Antioxidant Activity. PeerJ 2020, 8, e9769. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Mumper, R.J. Plant Phenolics: Extraction, Analysis and Their Antioxidant and Anticancer Properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Viacava, G.E.; Goyeneche, R.; Goñi, M.G.; Roura, S.I.; Agüero, M.V. Natural Elicitors as Preharvest Treatments to Improve Postharvest Quality of Butterhead Lettuce. Sci. Hortic. 2018, 228, 145–152. [Google Scholar] [CrossRef]
- Nguyen, T.T.H.; Cho, S.O.; Ban, J.Y.; Kim, J.Y.; Ju, H.S.; Koh, S.B.; Song, K.-S.; Seong, Y.H. Neuroprotective Effect of Sanguisorbae Radix against Oxidative Stress-Induced Brain Damage: In Vitro and in Vivo. Biol. Pharm. Bull. 2008, 31, 2028–2035. [Google Scholar] [CrossRef] [PubMed]
- Ginovyan, M.; Petrosyan, M.; Trchounian, A. Antimicrobial Activity of Some Plant Materials Used in Armenian Traditional Medicine. BMC Complement. Altern. Med. 2017, 17, 50. [Google Scholar] [CrossRef] [PubMed]
- Gatto, M.A.; Sanzani, S.M.; Tardia, P.; Linsalata, V.; Pieralice, M.; Sergio, L.; Venere, D.D. Antifungal Activity of Total and Fractionated Phenolic Extracts from Two Wild Edible Herbs. NS 2013, 05, 895–902. [Google Scholar] [CrossRef]
- Abad, M.J.; Guerra, J.A.; Bermejo, P.; Irurzun, A.; Carrasco, L. Search for Antiviral Activity in Higher Plant Extracts. Phytother. Res. 2000, 14, 604–607. [Google Scholar] [CrossRef] [PubMed]
- Solowey, E.; Lichtenstein, M.; Sallon, S.; Paavilainen, H.; Solowey, E.; Lorberboum-Galski, H. Evaluating Medicinal Plants for Anticancer Activity. Sci. World J. 2014, 2014, 721402. [Google Scholar] [CrossRef]
- Memete, A.R.; Timar, A.V.; Vuscan, A.N.; Miere (Groza), F.; Venter, A.C.; Vicas, S.I. Phytochemical Composition of Different Botanical Parts of Morus Species, Health Benefits and Application in Food Industry. Plants 2022, 11, 152. [Google Scholar] [CrossRef]
- Mirzavi, F.; Rajabian, A.; Boroumand-Noughabi, S.; Hosseini, A.; Boroushaki, M.T.; Hassanzadeh, S. Standardized Extract of Sanguisorba Minor Attenuates Injury in Aging Rat Model via the Nrf2/HO-1 Pathway. Acta NeuroBiol. Exp. (Wars) 2022, 82, 433–441. [Google Scholar] [CrossRef]
- Akbari, S.; Soodi, M.; Hajimehdipoor, H.; Ataei, N. Protective Effects of Sanguisorba Minor and Ferulago Angulata Total Extracts against Beta-Amyloid Induced Cytotoxicity and Oxidative Stress in Cultured Cerebellar Granule Neurons. J. Herbmed. Pharm. 2019, 8, 248–255. [Google Scholar] [CrossRef]
- Ansari, L.; Mahdinezhad, M.R.; Rakhshandeh, H.; Hosseini, A.; Noughabi, S.B.; Gholami, N.; Rajabian, A. Acute and Sub-Acute Toxicity Assessment of the Standardized Extract of Sanguisorba Minor. in Vivo. J. Complement. Integr. Med. 2022, 19, 987–997. [Google Scholar] [CrossRef]
- Terrab, A.; Valdés, B.; Díez, M.J. Study of Plants Visited by Honeybees (Apis Mellifera L.) in the Central Rif Region (N. Morocco) Using Pollen Analysis. Grana 2005, 44, 209–215. [Google Scholar] [CrossRef]
- Akassou, I.; Avosani, S.; Caorsi, V.; Verrastro, V.; Ciolli, M.; Mazzoni, V. Intrasexual Vibrational Behavior of Philaenus Spumarius in Semi-Field Conditions. Insects 2021, 12, 584. [Google Scholar] [CrossRef] [PubMed]
- Ganassi, S.; Domenico, C.D.; Altomare, C.; Samuels, G.J.; Grazioso, P.; Cillo, P.D.; Pietrantonio, L.; De Cristofaro, A. Potential of Fungi of the Genus Trichodermafor Biocontrol of Philaneus spumarius, the Insect Vector for the Quarantine Bacterium Xylella fastidosa. Pest Manag. Sci. 2023, 79, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Trkulja, V.; Tomić, A.; Iličić, R.; Nožinić, M.; Milovanović, T.P. Xylella Fastidiosa in Europe: From the Introduction to the Current Status. Plant Pathol J. 2022, 38, 551–571. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 | Sanguisorba minor subsp. balearica | [36] |
| 2 | Sanguisorba minor subsp. lasiocarpa | [37] |
| 3 | Sanguisorba minor subsp. magnolii | [38] |
| 4 | Sanguisorba minor subsp. mauritanica | [39] |
| 5 | Sanguisorba minor subsp. muricata | [40] |
| 6 | Sanguisorba minor subsp. verrucosa | [41] |
| S. minor Part | Extraction Solvent | Total Phenols | References |
|---|---|---|---|
| Roots | 96% Ethanol | 457.45 ± 4.59 μg GAE/mg | [25] |
| 70% Ethanol | 3.89 ± 0.01 mg GAE/g dw | [4] | |
| Aerial parts (stem + leaves) | Methanol | 132.80 ± 3.87 μg GAE/mg | [11] |
| Chloroform | 67.87 ± 0.77 μg GAE/mg | [11] | |
| Stem | 70% Ethanol | 0.18 ± 0.02 mg GAE/g dw | [4] |
| Leaves | 70% Ethanol | 1.19 ± 0.11 mg GAE/g dw | [4] |
| Compounds | Roots | Leaves | Flowers | Aerial Parts (Stems + Leaves) | References |
|---|---|---|---|---|---|
| Tannins | |||||
| 2,3-Hexahydroxydiphenoyl-glucose | 15.12 ± 0.21 | 11.59 ± 0.38 | 17.82 ± 0.60 | nd | [44] |
| Sanguiin H-10 derivative | 5.00 ± 0.06 | 1.75 ± 0.03 | 6.76 ± 0.02 | nd | [44] |
| 13.0 ± 0.30 | nd | nd | 5.14 ± 0.02 | [28] | |
| nd | nd | nd | 18.3 ± 0.40 | [59] | |
| 6.12 ± 0.05 | 9.82 ± 0.03 | nd | nd | [55] | |
| Punicalagin gallate | 28.58 ± 0.04 | 25.93± 0.02 | 18.67 ± 0.02 | nd | [44] |
| 21.7 ± 0.70 | nd | nd | nd | [28] | |
| 11.5 ± 0.20 | nd | nd | nd | [55] | |
| Sanguiin H-1 | 9.53 ± 0.02 | 13.46 ± 0.04 | 26.03 ± 0.05 | nd | [44] |
| Galoyl-bis-hexahydroxydiphenyl –glucoside, isomer 1 | 5.11 ± 0.22 | nd | nd | nd | [44] |
| 11.1 ± 0.20 | nd | nd | nd | [28] | |
| nd | 5.5 ± 0.10 | [59] | |||
| 12.25 ± 0.02 | nd | nd | nd | [55] | |
| Galloyl-bis-hexahydroxydiphenyl-glucoside, isomer 2 | 4.28 ± 0.03 | nd | nd | nd | [44] |
| 13.13 ± 0.07 | nd | nd | nd | [28] | |
| Ellagic acid hexoside | nd | 11.49 ± 0.04 | 0.89 ± 0.03 | nd | [44] |
| nd | nd | nd | 3.8 ± 0.10 | [28] | |
| nd | 6.2 ± 0.10 | nd | nd | [55] | |
| Ellagic acid pentoside | 0.24 ± 0.02 | 1.16 ± 0.02 | 1.02 ± 0.05 | nd | [44] |
| 11.66 ± 0.04 | nd | nd | nd | [28] | |
| 5.47 ± 0.008 | nd | nd | [55] | ||
| Pedunculagin | 8.0 ± 0.20 | nd | nd | nd | [28] |
| nd | 10.2 ± 020 | nd | nd | [55] | |
| Lambertianin C | 92.9 ± 0.20 | nd | nd | 22.3 ± 0.30 | [28] |
| nd | nd | nd | 18.6 ± 0.30 | [59] | |
| nd | 9.82 ± 0.03 | nd | nd | [55] | |
| Flavonoids | |||||
| C-type (epi)catechin trimer | 6.27 ± 0.02 | nd | nd | nd | [44] |
| Cyanidin-glucoside | nd | nd | 0.13 ± 0.07 | nd | [44] |
| B-type (epi)catechin dimer, isomer 1 | 3.44 ± 0.03 | 13.38± 0.03 | 10.26 ± 0.02 | nd | [44] |
| 37.4 ± 0.90 | nd | nd | 15.4 ± 0.30 | [28] | |
| 8.2 ± 0.80 | nd | nd | nd | [55] | |
| Catechin | 8.58 ± 0.02 | 15.42 ± 0.02 | 6.42 ± 0.01 | nd | [44] |
| 28.0 ± 0.50 | nd | nd | 20.4 ± 0.10 | [28] | |
| Cyanidin-malonylglucoside | nd | nd | 0.06 ± 0.02 | nd | [44] |
| B-type (epi)catechin dimer, isomer 2 | 11.79 ± 0.05 | 9.77 ± 0.03 | 3.71 ± 0.03 | nd | [44] |
| 48.8 ± 0.30 | nd | nd | 16.69 ± 0.07 | [28] | |
| Quercetin-galloyl-glucoside | nd | 1.95 ± 0.02 | 1.08 ± 0.01 | nd | [44] |
| Quercetin-glucuronide | nd | 20.20 ± 0.02 | 8.33 ± 0.02 | nd | [44] |
| nd | nd | nd | 18.0 ± 0.10 | [59] | |
| nd | 7.6 ± 0.20 | nd | nd | [55] | |
| nd | nd | 9.31 ± 0.05 | [28] | ||
| Quercetin-glucoside | nd | 8.17 ± 0.09 | 8.61 ± 0.04 | nd | [44] |
| Quercetin-galloylhexoside | nd | nd | nd | 1.320 ± 0.001 | [28] |
| nd | 0.842 ± 0.04 | nd | nd | [55] | |
| Quercetin-O-hexoside gallate (isomer 1) | nd | nd | nd | 4.5 ± 0.10 | [59] |
| Quercetin-O-hexoside gallate (isomer 2) | nd | nd | nd | 5.3 ± 0.20 | [59] |
| Quercetin-O-pentoside | nd | nd | nd | 1.521 ± 0.007 | [28] |
| Kaempferol-glucuronide | nd | 6.26 ± 0.02 | 2.12 ± 0.09 | nd | [44] |
| nd | 0.944 ± 0.02 | nd | nd | [55] | |
| Kaempferol-3-O-glucoside | nd | nd | nd | 11.3 ± 0.60 | [59] |
| Kaempferol-O-hexoside | nd | nd | nd | 9.84 ± 0.04 | [59] |
| Apigenin-O-deoxyhexoside | nd | nd | nd | 10.54 ± 0.01 | [28] |
| Phenolic acids | |||||
| 3-Caffeoylquinic acid (Neochlorogenic acid) | 3.42 ± 0.02 | 3.79 ± 0.04 | 2.11 ± 0.02 | nd | [44] |
| nd | nd | nd | 22 ± 1.00 | [59] | |
| Caffeic acid-glucoside | 10.90 ± 0.02 | 4.73 ± 0.01 | 2.01 ± 0.02 | nd | [44] |
| 5-Caffeoylquinic acid (Chlorogenic acid) | 2.10 ± 0.03 | 3.73 ± 0.01 | 3.96 ± 0.05 | nd | [44] |
| p-Coumaroylquinic acid | 4.08 ± 0.07 | 8.09 ± 0.07 | 11.48 ± 0.03 | nd | [44] |
| nd | 6.05 ± 0.25 | nd | nd | [55] | |
| Gallic acid glucoside | nd | nd | nd | 26 ± 2.00 | [59] |
| Caffeoyl ester (isomer 1) | nd | nd | nd | 14.8 ± 0.10 | [59] |
| Digalloyl glucoside | nd | nd | nd | 22 ± 1 | [59] |
| 9.5 ± 0.10 | 7.4 ± 0.30 | nd | nd | [55] | |
| Caffeoyl ester (isomer 2) | nd | nd | nd | 5.7 ± 0.20 | [59] |
| Ellagic acid | 2.85 ± 0.03 | nd | nd | nd | [44] |
| 13.3 ± 0.40 | nd | nd | nd | [28] | |
| 4.3 ± 0.10 | nd | nd | nd | [55] | |
| Sample | Method | Result | References |
|---|---|---|---|
| Essential oil of S. minor aerial parts 0.1 mg/mL (Portugal) | DPPH | 11% of inhibition | [65] |
| Carotene-linoleic acid assay | 99% of inhibition | ||
| Ethanolic extract of S. minor aerial parts 0.1 mg/mL (Portugal) | DPPH | 93% of inhibition | |
| Decoction of S. minor aerial parts 0.1 mg/mL (Portugal) | DPPH | 93% of inhibition | |
| Carotene-linoleic acid assay | 95% of inhibition | ||
| Aqueous extract of S. minor leaves 5 mg/mL (Spain) | Peroxyl radical (H2O2) scavenging | 64.35% of inhibition | [56] |
| Hydroxyl radical (OH•) scavenging | 33.50% of inhibition | ||
| Water/ethanol extract of S. minor leaves 0.1–0.01 g/mL (Italy) | Peroxyl radical scavenging | 212 LOO•kg of fresh plant | [67] |
| FRAP | 257 mmol Fe2+/kg | ||
| Ethanolic extract of S. minor roots (Serbia) | ABTS | 77.54 µg TE/mg | [25] |
| DPPH | 96.51 µg TE/mg | ||
| CUPRAC | 346.49 µg TE/mg | ||
| FRAP | 188.22 µg Fe/mg | ||
| TRP | 1.16 µg AAE/mg | ||
| Methanolic extract of S. minor roots (Serbia) | ABTS | 76.97 µg TE/mg | |
| DPPH | 97.29 µg TE/mg | ||
| CUPRAC | 34.35 µg TE/mg | ||
| FRAP | 214.02 µg Fe/mg | ||
| TRP | 1.19 µg AAE/mg | ||
| Chloroform extract of S. minor roots (Serbia) | ABTS | 46.11 µg TE/mg | |
| DPPH | 40.51 µg TE/mg | ||
| CUPRAC | 96.80 µg TE/mg | ||
| FRAP | 48.02 µg Fe/mg | ||
| TRP | 0.11 µg AAE/mg | ||
| Methanolic extract of S. minor aerial parts (Serbia) | ABTS | 77.26 µg TE/mg | [11] |
| DPPH | 95.06 µg TE/mg | ||
| CUPRAC | 289.09 µg TE/mg | ||
| FRAP | 205.62 µg Fe/mg | ||
| TRP | 0.58 µg AAE/mg | ||
| Chloroform extract of S. minor aerial parts (Serbia) | ABTS | 53.83 µg TE/mg | |
| DPPH | 40.31 µg TE/mg | ||
| CUPRAC | 182.90 µg TE/mg | ||
| FRAP | 78.22 µg Fe/mg | ||
| TRP | 0.06 µg AAE/mg | ||
| Ethanolic extract of S. minor roots (Romania) | DPPH | 92.93% of inhibition | [4] |
| FRAP | 10.81 μmol TE/g | ||
| Ethanolic extract of S. minor stems (Romania) | DPPH | 0.32% of inhibition | |
| FRAP | 0.16 μmol TE/g | ||
| Ethanolic extract of S. minor leaves (Romania) | DPPH | 43.15% of inhibition | |
| FRAP | 2.88 μmol TE/g |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tocai, A.C.; Kokeric, T.; Tripon, S.; Barbu-Tudoran, L.; Barjaktarevic, A.; Cupara, S.; Vicas, S.I. Sanguisorba minor Scop.: An Overview of Its Phytochemistry and Biological Effects. Plants 2023, 12, 2128. https://doi.org/10.3390/plants12112128
Tocai AC, Kokeric T, Tripon S, Barbu-Tudoran L, Barjaktarevic A, Cupara S, Vicas SI. Sanguisorba minor Scop.: An Overview of Its Phytochemistry and Biological Effects. Plants. 2023; 12(11):2128. https://doi.org/10.3390/plants12112128
Chicago/Turabian StyleTocai (Moţoc), Alexandra Cristina, Tijana Kokeric, Septimiu Tripon, Lucian Barbu-Tudoran, Ana Barjaktarevic, Snezana Cupara, and Simona Ioana Vicas. 2023. "Sanguisorba minor Scop.: An Overview of Its Phytochemistry and Biological Effects" Plants 12, no. 11: 2128. https://doi.org/10.3390/plants12112128